

On Feb 25, 2022, a paper entitled “Distinct networks coupled with parietal cortex for spatial representations inside and outside the visual field” was online published in NeuroImage by Dr. Yuji Naya group in the School of Psychological and Cognitive Sciences at Peking University and the PKU-IDG/McGovern Institute for Brain Research.
While planning to reach a target, it is necessary to obtain its location information from “egocentric” spatial coordinates (Stein, 1989), which includes the space inside the visual field as well as the space outside the visual field. Nevertheless, previous studies mostly examined the target representation within the visual field (Abrahams et al., 1997; Wolbers et al., 2008; Guterstam et al., 2015), the neural representaiton of target outside the visual field has been neglected. Classic phenomenon, known as the “alignment effect” (Presson and Hazelrigg, 1984) or “front facilitation” (Kelly and McNamara, 2009), suggested that the external spaces surrounding our body might be coded heterogeneously by different neural systems depending on the target location along the anterior-posterior axis of the self-body. For example, a decreased performance (Attneave and Farrar, 1977) or prolonged response latency (Sholl, 1987) was observed when locating a target behind self-body. The present study thus investigated the neural architectures that support the mental representations of egocentric space inside and outside the visual field.
fMRI and MEG were applied to a spatial-memory paradigm using a virtual environment for human participants (Fig. 1a)(Zhang and Naya, 2020). In the paradigm, participants walked towards three objects and stopped at the center of them (walking period), during which participants encoded the spatial relationship of the three objects. One of the three objects was then presented with the environmental background, which prompted participants to feel as if they were facing the objects in the virtual environment (facing period). Next, one of the other two objects was presented as the target object (targeting period), participants remembered the location of the target object relative to the self-body, this to-be-remembered targeting location could be in the left, right, or back positions relative to participants (Fig. 1b). This spatial-memory paradigm allowed us to compare the neural representations of egocentric space inside (left/right-target condition) and outside (back-target condition) of the visual field, and localize the target representation with a control condition, where no spatial demands were required in the facing and targeting period.
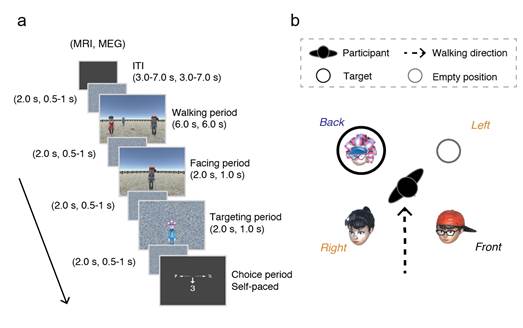
Fig. 1 Spatial-memory paradigm. (a) Each trial consisted of four periods. Walking period: participants walked toward three human characters using the first-person perspective and stopped on a wood plate in the center. Facing period: one of the human characters was presented, indicating the participant’s self-orientation. Targeting period: a photo of the target character was presented on a scrambled background. Choice period: participants chose the number on response cue indicating the position of the target character relative to their body. (b) The spatial relationship between the participant and the human characters corresponding to the examplar trial in (a), where the target was behind the participant (black circle).
Both fMRI and MEG experiments revealed that the frontoparietal network represented a retrieved target inside the visual field (Fig.2a,b,e,g). In contrast, the MTL-parietal network represented a retrieved target outside the visual field more although it was also involved in retrieving the target inside the visual field (Fig.2c,d,f,g). A high temporal-resolution of the MEG data showed that the activation of MTL-parietal network preceded the frontoparietal network during recall of the target location (Fig.2b,d,g). These findings, consistent with the “front facilitation” effect, exhibited disproportional spatial representations around the self-body, with a bias toward the left/right-target location in comparison with the back-target location. The parietal cortex may represent the present entire space around the self-body by interacting with the MTL to look back on the past as well as the dorsal-frontal regions to prepare for a subsequent action.
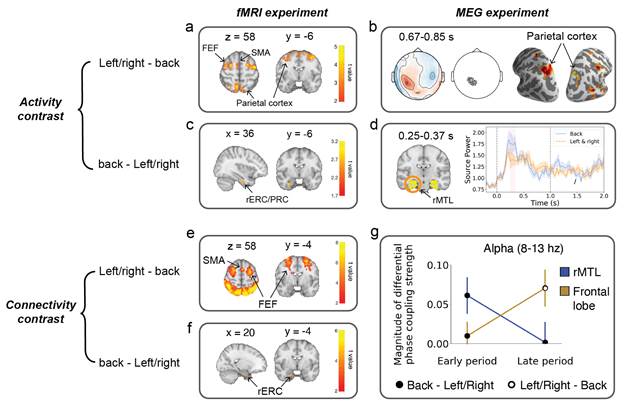
Fig. 2 The main results revealed from fMRI and MEG experiments. (a, b) larger activation was observed in the dorsal brain regions under the left/right-target conditions than the back-target condition. (c, d) larger activation was observed in the ventral brain regions (i.e. right ERC) for the back-target conditions relative to the left/right-target condition. (e, f) connectivity results are consistent with the results of activity contrast (a, b, c, d). (f) Alpha band phase coupling strength of the parietal cortex with FEF and SMA (frontoparietal network, yellow line) and with the ERC and PRC (MTL-parietal network, blue line) in early (0.25-0.37s) and late (0.67-0.85s) period.
Article link: https://www.sciencedirect.com/science/article/pii/S1053811922001707
This work was supported by the National Natural Science Foundation of China Grant (31421003), by the Fundamental Research Funds for the Central Universities, PKU (7100602954) and by the Innovations of Science and Technology 2030, China (2021ZD0203600). The first author of the present study is Bo Zhang, who obtained a Ph.D from the School of Psychological and Cognitive Sciences at Peking University, and now works as a joint Post-doc program in Beijing Academy of Artificial Intelligence and Tsinghua University. The corresponding author is Dr. Yuji Naya of Peking University.
Reference
Stein, J. F. (1989). Representation of egocentric space in the posterior parietal cortex. Q J Exp Physiol, 74(5), 583-606. https://doi.org/10.1113/expphysiol.1989.sp003314
Abrahams, S., Pickering, A., Polkey, C. E., & Morris, R. G. (1997). Spatial memory deficits in patients with unilateral damage to the right hippocampal formation. Neuropsychologia, 35(1), 11-24. https://doi.org/10.1016/s0028-3932(96)00051-6
Wolbers, T., Hegarty, M., Buchel, C., & Loomis, J. M. (2008). Spatial updating: how the brain keeps track of changing object locations during observer motion. Nat Neurosci, 11(10), 1223-1230. https://doi.org/10.1038/nn.2189
Guterstam, A., Bjornsdotter, M., Gentile, G., & Ehrsson, H. H. (2015). Posterior cingulate cortex integrates the senses of self-location and body ownership. Curr Biol, 25(11), 1416-1425. https://doi.org/10.1016/j.cub.2015.03.059
Presson, C. C., & Hazelrigg, M. D. (1984). Building spatial representations through primary and secondary learning. J Exp Psychol Learn Mem Cogn, 10(4), 716-722. https://doi.org/10.1037//0278-7393.10.4.716
Kelly, J. W., & McNamara, T. P. (2009). Facilitated pointing to remembered objects in front: evidence for egocentric retrieval or for spatial priming? Psychon Bull Rev, 16(2), 295-300. https://doi.org/10.3758/PBR.16.2.295
Attneave, F., & Farrar, P. (1977). The visual world behind the head. Am J Psychol, 90(4), 549-563. https://www.ncbi.nlm.nih.gov/pubmed/610446
Sholl, M. J. (1987). Cognitive maps as orienting schemata. J Exp Psychol Learn Mem Cogn, 13(4), 615-628. https://doi.org/10.1037//0278-7393.13.4.615
Zhang, B., & Naya, Y. (2020). Medial Prefrontal Cortex Represents the Object-Based Cognitive Map When Remembering an Egocentric Target Location. Cereb Cortex, 30(10), 5356-5371. https://doi.org/10.1093/cercor/bhaa117
